Il Passero del Giappone
Il
Passero del Giappone è uccello che si presta più di ogni
altro ad essere impiegato nell'allevamento degli Astrildidi ed i giapponesi
furono i primi ad utilizzarlo a questo fine, seguiti ben presto dagli europei.
Come
è noto, nel
Lo
studio del comportamento animale era ancora agli inizi ed i prodotti alimentari
disponibili sul mercato non avevano raggiunto l'elevato standard attuale, per
cui si fu obbligati a ricorrere a qualsiasi mezzo per riprodurre specie
ornitiche delicate e rare.
Intervenne
a questo punto il Passero del Giappone che, prodotto di reincroci, non
è in possesso di specifiche caratteristiche etologiche e non ha espresso
caratteristiche semantiche originali nei propri pulii pur avendo (reincrocio
sin che si vuole, ma è un uccello) assunto dalle forme parentali
ancestrali il carattere appreso dell'accettazione e del riconoscimento della
prole in funzione del luogo in cui questa si trova, e cioè il nido (per
la discussione, cfr. TRUFFI, 1980).
Si
adatta così con estrema facilità a covare le uova e ad allevare i
piccoli di tutte le specie di Astrildidi a becco grosso ed anche alcune di
quelle a becco sottile che pure gli sono filogeneticamente meno vicine.
|
|

Unica
condizione per il positivo esito dello svezzamento è che si tratti di
uccelli che nei primi giorni di vita non richiedano tassativamente
un'alimentazione strettamente insettivora. Anche in questo caso però il
risultato può non essere compromesso se si dispone di soggetti
preventivamente abituati ad alimentarsi almeno in parte con prede vive, che
somministreranno così ai nidiacei loro affidati.
Decisivo,
perché l'allevamento abbia luogo, è l'impiego di Passeri del
Giappone che abbiano uova o piccoli allo stesso stadio di sviluppo di
quelli dei quali devono prendersi cura.
E'
però consigliabile sostituire le loro uova e trasferire ad altri i loro
piccoli ad evitare che la stessa coppia debba dedicarsi contemporaneamente
all'allevamento dei propri nidiacei (che sarebbero immancabilmente meglio
alimentati ed accuditi, almeno in linea di larga massima) e di quelli di altre
specie. Da quando, l'allevamento dei diamanti australiani si è rivelato
economicamente positivo, le coppie di riproduttori vengono isolate ed
alloggiate in gabbie da cova (a volte anche piuttosto anguste) ponendo loro a
disposizione nidi rudimentali in cui le femmine depongono uova che vengono
regolarmente sottratte e passate alle balie per la cova e lo svezzamento,
procedimento che provoca una ovideposizione più numerosa di quanto non
si verificherebbe se i genitori naturali dovessero accudire alla propria
nidiata.
|
|

Si
tratta però di una selezione totalmente negativa perché
innesca un processo paragonabile a quanto avviene nell'allevamento dei polli o
delle quaglie condotto su scala industriale.
I
piccoli Astrildidi appena sgusciati dall'uovo vengono imprintizzati
dalla specie che vedono per prima (e questo irreversibile processo si verifica
in un breve spazio di tempo) e che di loro si occupa; in natura quindi dai
propri genitori, nella situazione che stiamo esaminando dai Passeri del
Giappone. Nel corso dell'allevamento le cose procedono a tal punto che i
giovani svezzati da questi ultimi saranno in seguito incapaci di accoppiarsi
con i propri conspecifici, manifestando una netta tendenza preferenziale a
scegliere i Passeri del Giappone (con i quali per effetto
dell'imprintizzaziane finiscono per identificarsi) come partners sessuali.
E'
questo uno dei motivi per cui negli allevamenti amatoriali di uccelli esotici
si incontrano facilmente tanti ibridi con il Passero del Giappone,
dovuti il più delle volte al caso.
|
|

Le
balie perciò, ai fini di un corretto allevamento, vanno usate solo in
casi estremi e quando non esiste nessun'altra possibilità, per
sopravvenuti imprevisti motivi, di salvare uova o nidiacei ed anche in questo
caso occorre poi drasticamente isolare i giovani non appena raggiunta
l'indipendenza ed allontanare dalla loro vista i genitori adottivi a
svezzamento avvenuto.
E'
chiaro che le positive caratteristiche di domesticità evidenziate hanno
fatto del Passero del Giappone un soggetto ideale per l'ibridazione
amatoriale. Quasi sempre sono state impiegate le femmine, proclivi alla
deposizione senza difficoltà alcuna, accoppiate a maschi di altre
specie, tutte però appartenenti alla famiglia degli Astrildidi.
Le
vecchie notizie risalenti ad oltre cinquant'anni fa (e mai più
riconfermate) relative ad avvenute ibridazioni con il Canarino domestico
e forse altri uccelli del genere Serinus appaiono in verità scarsamente
credibili.
Elencare
le ibridazioni ottenute di cui esiste traccia nella letteratura (non in tutti i
casi affidabile) non è cosa semplice e fatalmente si corre il rischio di
presentare dati incompleti oppure di prestare a certe notificazioni,
necessitanti di verifica, più credibilità di quanto nella
realtà non meritino.
Praticata
anche negli allevamenti italiani, tanto è vero che alle esposizioni
vengono presentati F 1 di notevoli caratteristiche che giustamente ottengono
lusinghieri riconoscimenti, l'ibridazione del Passero del Giappone si
è sviluppata in tre differenti direzioni.
La
prima, con precisi intendimenti scientifici e sistematici (scuola
tedesca, e molto dobbiamo in questo senso al Russ, che se ne occupò alla
fine del secolo scorso) per stabilirne le origini.
La
seconda (scuola tedesca in un primo tempo, successivamente olandese e
danese ed ultimamente ancora tedesca) volta alla ricostruzione di soggetti
monocolori scuri e con nette squamature sulle parti inferiori allineate e
precise come previsto dai più rigorosi standards di eccellenza, anche se
forse attualmente un po' lontani dallee realtà di allevamento tanto
è vero che i criteri di giudizio tedeschi prevedono in ogni categoria
sia soggetti con disegno sia a ventre chiaro e non disegnato.
La
terza, di scuola latina (se così si può affermare, dato
che assolutamente nulla di concreto è stato fatto se non lasciando che
il caso si sbizzarrisse a suo piacere) si è sviluppata senza alcuna
finalizzazione razionale e tendendo a produrre ibridi ai soli fini espositivi,
appaganti sotto il profilo estetico, giudicati in relazione alla maggiore o
minore estrinsecazione delle caratteristiche delle specie che hanno contribuito
alla loro realizzazione.
Analizziamole in dettaglio.
Allevato
dai giapponesi sin dagli inizi del 1700 (e in un tempo relativamente breve
arrivarono a selezionarne oltre quaranta varietà, tutte abbastanza
stabilmente fissate) e di probabile provenienza cinese, il Passero del
Giappone fece la sua prima comparsa in Europa ad una esposizione inglese
nel 1860. Successivamente il Russ ne importò alcune coppie
in Germania negli ultimi anni dell'ottocento e ne curò l'allevamento,
rendendolo se non popolare almeno abbastanza conosciuto con il pubblicarne note
pertinenti su una rivista amatoríale da lui diretta.
Per
lungo tempo si è ritenuto che all'origine avessero contribuito incroci
fra specie e sottospecie asiatiche e persino africane, quali il Domino Lonchura
punctulata ed il Becco d'argento Lonchura malabarica cantans,
dato che questi uccelli sotto certi aspetti presentano caratteri fenotipici di
disegno e colore che richiamano abbastanza da vicino quelli del Passero
del Giappone. Da qui la tesi ancora oggi diffusa di " uccello
artificiale ".
|
|
|
|
Lonchura malabarica cantans |
Lonchura punctulata |


La
letteratura è invece attualmente concorde nell'affermarne la diretta
discendénza dal Cappuccino a coda lunga Lonchura striata,
anch'esso da lungo tempo presente allo stato captivo negli allevamenti
amatoriali asiatici.
|
|
|
Lonchura striata |

Il
Cappuccino a coda lunga presenta un'ampia suddivisione in sottospecie e
razze geografiche, che hanno tra loro differenze più o meno marcate. Le
stesse variazioni si riscontrano nel disegno scuro del Passero del Giappone
e non furono inizialmente notate forse perché nei primi ottant'anni del
suo allevamento come uccello da gabbia tutte le attenzioni erano dedicate a
produrre soggetti pezzati di bianco. Con la domesticazione tutti gli animali
manifestano prima o poi uno schiarimento del pelo o delle penne perché
non necessitano più di protezione mimetica in quanto già protetti
dall'uomo.
Originariamente
si ebbero solo alcune parti del piumaggio di colore bianco che a seguito
dell'allevamento di selezione si estesero sino allo schiarimento totale con
tutte le gradazioni intermedie, da una preponderanza delle zone di colore scuro
ad una preponderanza di quelle chiare. Per consentirci di identificare
l'effettiva derivazione e collocazione di questi Astrildidi domestici
è necessario fare riferimento ad alcune differenze di certe razze
originarie di Lonchura striata, i cui areali, ricavati da ALI e RIPLEY
(1974) e da CHENG ( 1976 ) sono riportati in cartina.
|
|
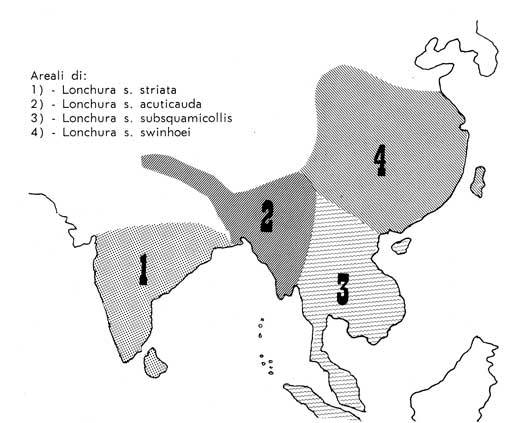
La
forma tipo, Lonchura striata striata (Linneo 1776), è
superiormente, compreso testa e collo, di colore quasi esclusivamente nero
bruno mentre inferiormente presenta un crema luminoso che a distanza può
apparire quasi bianco.
Esistono
varie sottospecie, di cui alcune endemícamente insulari, come Lonchura
striata fumigata (Walden 1873) delle isole Andamans e Lonchura striata
semistriata (Huma 1874) delle isole Nicobar, con il groppone scuro ed
altre, tutte continentali, con il groppone chiaro, come Lonchura
striata acuticauda (Hodgson 1836) in cui il piumaggio nero bruno del dorso
viene interrotto solo scarsamente da piccole striscioline chiare longitudinali.
Forse per il disegno, ma più sicuramente per ragioni geografiche, le
forme subspecifiche con il groppone scuro parrebbero a mio avviso non essere
state usate dai cinesi per la selezione del Passero del Giappone, ma piuttosto,
oltre a Lonchura striata acuticauda, anche Lonchura striata
subsquamicollis (Stuart Baker 1925) il cui areale di diffusione confina con
Sono
entrambe di colore cioccolato, soprattutto al collo, e presentano su tutto
il piumaggio una maggiore striatura. Le parti inferiori sono decisamente grigio
bruno. In complesso sono di colori più tenui ed il groppone è
spruzzato di grigio. L'aspetto generale è piuttosto monocorde e Lonchura
striata subsquamicollis si differenzia da Lonchura striata swinhoei
per il groppone ancora più chiaro, simile a quello di Lonchuraa
striata acuticauda.
Queste
differenze di disegno e di intensità di colore ritornano nella semi
totalità dei casi nella maggior parte dei Passeri del Giappone e
si possono ancora ritrovare abbastanza riconoscibili nei soggetti pezzati di
chiaro. Pezzati più scuri mostrano al collo piume brune più
chiare e sono generalmente molto chiari sull'alto petto e nella parte bassa del
collo. E' ancora sufficientemente distinguibile una striatura diritta grigio
bruna.
Se
invece i Passerí del Giappone fossero derivati (cosa che a mio
avviso non è) dalla forma tipo, le zone attorno alla gola e sul groppone
avrebbero dovuto essere prevalentemente bianche ed il groppone anche scuro
monocolore, come abbiamo ricordato in precedenza accennando alle varie forme
subspecifiche. Il groppone dei pezzati con colore di base scuro è di
solito lavato di grigio.
Si
può ragionevolmente quindi concludere che gli antenati ancestrali si
limitino (più che a Lonchura striata acuticauda ) a Lonchura
striata subsquamicollis e Lonchura striata swinhoei, mentre invece
tutte le deviazioni di colore o disegno sono da ricondursi a mutazioni
spontanee oppure ad incroci con altre specie, che ancora oggi esercitano nella
discendenza la loro influenza sulle caratteristiche del fenotípo.
Nei
primi decenni dell'allevamento del Passero del Giappone la tendenza dei
selettori era orientata a far riprodurre i soggetti importati dal Giappone
cercando di ottenere sfumature di colore diverse e più chiare. Si ebbe
così la creazione di varietà ormai ben codificate, dal bruno al
giallastro al crema al nero diluito ai pezzati, sino al bianco.
Successivamente
si è venuto da anni delineando l'orientamento ad eliminare nuovamente il
bianco delle pezzature e la diluizione di colore per ritornare a soggetti
completamente scuri. Su base selettiva la cosa però si presenta difficile,
dato che si tratta di invertire la tendenza (dopo un secolo di allevamento) che
aveva prodotto uccelli in cui il bianco (più o meno esteso e regolare)
era quasi sempre presente. In un primo tempo si pensò di procedere, per
il " reincrocio all'indietro ", con le diverse sottospecie di
partenza ma ben presto questa strada venne abbandonata dopo aver constatato che
nelle varie razze di Lonchura striata, e soprattutto con Lonchura
striata acuticauda di cui sul mercato era disponibile e a volte lo è
tuttora un'offerta piuttosto ampia, si osservava nell'allevamento in purezza
allo stato captivo la comparsa di una pezzatura bianca che iniziando dalla
radice del becco si estendeva successivamente in tempi brevissimi (una o due
generazioni) alle rimanenti parti del piumaggio.
Fu
ritenuto quindi opportuno, ed in parecchi casi positivamente gratificante,
l'introdurre sangue nuovo procedendo in direzione di accoppiamenti con specie
filogeneticamente vicine (per un esame più approfondito cfr. RADTKE
1979). Senza entrare in questa sede in una analisi tecnica dettagliata dei
risultati ottenuti e delle verifiche da questi scaturite, argomenti che
dovranno comunque essere oggetto di discussione sulla stampa specializzata e
che prima o poi bisognerà affrontare per uscire dal generico e
sviluppare, acquisendo nel nostro bagaglio di cognizioni le esperienze
conseguite all'estero, un corretto lavoro finalizzato di specializzazione, mi
limiterò ad accennare che gli ibridi ottenuti ai fini della
ricostruzione della tipicità non presentano, all'occhio del profano,
né pregi né caratteristiche particolari ed in sede espositiva
sono scarsamente interessanti ( se il giudicante è padrone della materia
li valorizzerà nelle voci appropriate della scheda) mentre invece
possono costituire materiale validissimo in allevamento per il miglioramento
dei ceppi, anche se la fertilità in F 1 riscontrata ( ma assolutamente
non generalizzata) nei maschi è pressoché nulla nelle femmine ed
è quindi giocoforza continuare l'allevamento in R 1, accoppiando i
maschi con Passeri del Giappone femmina o, più raramente, con
femmine della specie parentale.
Gli
R i presentano buona fertilità anche nelle femmine ed è quindi
possibile procedere ad accoppiamenti compensativi e meglio indicizzati
soprattutto per quanto si riferisce al tipo, dato che gli ibridi ottenuti
possono a volte essere migliorativi ai fini del colore e del disegno ma quasi
sempre presenteranno caratteristiche debordanti per la sagoma, troppo
affusolata in alcuni casi e troppo raccolta ed arrotondata in altri, e per la
posizione assunta, quasi sempre eccessivamente eretta sul posatoio. A
titolo di informazione ricordo che i giudici olandesi considerano non fra gli
ibridi ma come Passeri del Giappone i risultati dell'accoppiamento con
le varie forme di Lonchura striata (VAN DEN MOLEN 1971) .
Gli
R 2 e gli R 3 sono totalmente fecondi in entrambi i sessi.
Le
ibridazioni conseguite ai fini sopra accennati sono state ottenute per lo
più con specie asiatiche del genere Lonchura (i Cappuccini) e
precisamente (almeno per quanto a mia conoscenza) con: Cappuccino del Borneo
Lonchura fuscans, Cappuccino a ventre ondulato Lonchura
molucca, Cappuccino montano Lonchura kelaarti, Cappuccino
di Giava, Lonchura leucogastroides, Cappuccino triste Lonchura
tristis, Cappuccino a testa bianca Lonchura maja, Cappuccino
a testa nera Lonchura malacca, Cappuccino tricolore Lonchura
malacca atricapilla, Domino Lonchura punctulata.
|
|
|
|
Lonchura fuscans |
Lonchura leucogastroides |


|
|
|
|
Lonchura punctulata |
Lonchura molucca |

|
|
|
|
Lonchura maja |
|


Interessanti
esperimenti che hanno dato origine anche ad ibridi esteticamente pregevoli e
dai quali in un primo tempo sembrava di poter ottenere risultati eccezionali
(calda colorazione bruno intenso sulle parti superiori con mascherina nerastra,
groppone nero intenso, parti inferiori più o meno perlate di grigio o
squamate su fondo scuro, soggetti lievemente più grossi dei Passeri
del Giappone, con portamento eretto e coda leggermente più corta e
meno lanceolata) si sono ottenuti dall'accoppiamento con il Donacola a petto
castano Lonchura castaneothorax.
Purtroppo,
non fu possibile continuarne l'allevamento data la quasi totale
sterilità delle femmine anche nei reincroci di seconda e terza
generazione ( RADTE 1989).
Poco
interessanti, poiché di taglia ridotta, gli ibridi con
|
|
|
|
Lonchura bicolor |
Lonchura malabarica |


Di non spiccate caratteristiche anche gli
ibridi ottenuti con il Becco d'argento Lonchura malabarica cantans
e con il Becco di piombo Lonchura malabarica malabarica, interessanti
però per l'armonica fusione delle sfumature di colore della livrea,
disturbata da una più o meno vistosamente presente tacca gulare
bíanchíccia bordata di nerastro, ed a mio parere più che
accettabili per quanto si riferisce al tipo.
Ai soli fini
espositivi poi si sono avuti incroci con varie specie, per lo più di
origine australiana, e precisamente con il Diamante a coda lunga Poephila
acuticauda, il Diamante a bavetta Poephila cincta, il Diamante
mascherato Poephila personata, il Diamante mandarino Taeniopygia
guttata castanotis, il Diamante a coda rossa Neochmia
ruficauda, il Diamante di Bicheno Poephila bichenovii, il Diamante
guttato Emblema guttata (a proposito di quest'ultima ibridazione, a
mio ricordo mai presentata in una esposizione italiana, cfr. Buchan 1991),
alcuni di questi di veramente appaganti caratteristiche estetiche e che sino ad
ora non hanno offerto casi di fertilità, manifestatisi invece (ma
raramente e per quanto mi consta solo nei maschi) con il Padda Padda
oryxívora di cui sono state utilizzate sia la forma grigia ancestrale
sia le varietà bianca e isabella.